
Efecto de diversas láminas de riego sobre las etapas fenológicas del cultivo de tomate (Solanum lycopersicum L.), en municipio de Totogalpa, corredor seco de Nicaragua
Effect of different irrigation sheets on the phenological stages of the tomato crop (Solanum lycopersicum L.), in the municipality of Totogalpa, dry corridor of Nicaragua
Javier Antonio Barberena Moncada
Universidad Nacional Autónoma de Nicaragua. Centro para la Investigación en Recursos Acuáticos de Nicaragua. CIRA/UNAN-Managua.
https://orcid.org/0000-0001-9119-4485
javier.barberena@cira.unan.edu.ni
Recibido
21/06/2023
Aceptado
16/11/2023
RESUMEN
El calentamiento global ha provocado que se busquen alternativas genéticas para hacer los cultivos resistentes a condiciones secas y temperaturas extremas, también se buscan métodos para suplir la demanda hídrica de los cultivos en época seca o en inviernos con reducidas precipitaciones. Técnicas como los sistemas de riego por goteo han solucionado la necesidad hídrica de diversos cultivos, maximizando el consumo de agua, y reduciendo las perdidas por escorrentía. La presente investigación se basó en evaluar el efecto de la aplicación de diferentes láminas de riego sobre las etapas fenológicas del cultivo de tomate (Solanum lycopersicum L.) en el municipio de Totogalpa, corredor seco de Nicaragua. Las láminas de aguas suplieron el 60% (T1), 80% (T2) y 100% (T3) de las pérdidas de agua por evapotranspiración en las plantas de tomate. Se utilizó un diseño completo al azar con tres repeticiones. el experimento fue al aire libre en Totogalpa, Nicaragua. Las etapas vegetativas y reproductivas de la planta de tomate fueron determinadas mediante modelo de tiempo térmicos (GDD). Pruebas de significancias al 0.05% fueron realizadas para determinar si había diferencias entre los tratamientos. Resultados obtenidos indican que la etapa vegetativa necesita 502 GDD a los 34 días después del trasplantes (DDT), con diferencias (p < 0.05) en las variables diámetro de tallo principal y número de hojas; la etapa reproductiva 1200 GDD y 77 DDT para generar la maduración y cosecha del fruto; y en la etapa reproductiva se encontraron diferencias (p< 0.05) entre los tratamientos en cuanto a número de flores/planta, número de frutos/planta, diámetro polar y ecuatorial de fruto y peso de fruto. Entre los tratamientos los mejores resultados se dan al suplir el 100% de pérdidas por evapotranspiración en las plantas de tomate, debido a que presentan los mejores promedios en la etapa vegetativa y reproductiva.
PALABRAS CLAVES
Déficit hídrico; estrés hídrico; fenología de tomate; láminas de riego; tiempo termal; uso eficiente de agua.
ABSTRACT
Global warming has caused the search for genetic alternatives to make crops resistant to dry conditions and extreme temperatures, and methods are also being sought to meet the water demand of crops in dry seasons or in winters with reduced rainfall. The present investigation was based on evaluating the effect of the application of different irrigation sheets on the phenological stages of the tomato crop (Solanum lycopersicum L.) in the municipality of Totogalpa, dry corridor of Nicaragua. The water sheets supplied 60% (T1), 80% (T2) and 100% (T3) of the water losses by evapotranspiration in tomato plants. A complete randomized design with three replications was used. The experiment was conducted outdoors in Totogalpa, Nicaragua. The vegetative and reproductive stages of the tomato plant were determined using a thermal time model (TDM). Significance tests at 0.05% were performed to determine if there were differences between treatments. Results obtained indicate that the vegetative stage requires 502 GDD at 34 days after transplanting (DDT), with differences (p < 0.05) in the variables main stem diameter and number of leaves; the reproductive stage requires 1200 GDD and 77 DDT to generate fruit ripening and harvest; and in the reproductive stage, differences (p < 0.05) were found among treatments in terms of number of flowers/plant, number of fruits/plant, polar and equatorial fruit diameter and fruit weight. Among the treatments, the best results were obtained by supplying 100% of the losses due to evapotranspiration in tomato plants, due to the fact that they presented the best averages in the vegetative and reproductive stages.
KEYWORDS
Water deficit; water stress; tomato phenology; irrigation sheets; thermal time; efficient water use.
INTRODUCCIÓN
Cerca de 33947 km2 (26%) del territorio de Nicaragua es parte de lo que se denomina Corredor Seco Centroamericano, una región que abarca desde Chiapas, México, la zona central de Guatemala, Honduras y Nicaragua, zonas bajas de El Salvador, hasta culminar en Guanacaste (Costa Rica), incluye el arco seco de Panamá (Fraga, 2020; van der Zee Arias et al., 2012). El corredor seco se caracteriza por tener condiciones climatológicas muy distintas Fraga (2020) y van der Zee Arias (2012), que varían según la zona climática y productiva, generando tres zonas con intensidades de sequías. Zonas con sequías severas, presentan régimen de precipitación entre 800-1200 mm/año, con seis meses secos y evaporación potencial de 200 mm/mes.
Zonas con sequías altas, presentan regímenes pluviales 1200-1600 mm/año, con cuatro a seis meses secos y evaporación potencial 130 mm/mes. Zonas con sequía bajas, con precipitaciones superiores a 1600-2000 mm/año, cuatro a seis meses secos, evapotranspiración menor a 100 mm/mes. También, se suman los fenómenos climáticos como sequías prolongas producidas por el Niño y la Oscilación del Sur (ENOS) y la distribución anómala de las precipitaciones dentro del periodo lluvioso que es conocido como veranillo de invierno o canícula (Magaña et al., 1999).
La variabilidad climática tiene como una de sus principales consecuencias la afectación de la seguridad alimentaria, disminuyendo el rendimiento de los cultivos (IPCC, 2019). En la sequía del año 2015, se vieron impactados los departamentos de Carazo, Chinandega, Boaco, Estelí, Madriz y Nueva Segovia, donde las pérdidas en los cultivos representaron el 50% a nivel departamental y el 30% de la producción nacional (Calvo-Solano et al., 2018).
El cultivo de tomate es la segunda hortaliza de mayor importancia a nivel mundial (Jiménez-Martínez et al., 2015). En Nicaragua también es la segunda hortaliza, pero en cuanto a consumo y comercialización, llegando a producirse hasta 2469 hectáreas (Duarte-Canales & Benavides González, 2020; Jiménez-Martínez et al., 2015). El tomate es sensible a los cambios en el régimen climático (temperatura, radiación solar y agua disponible en el suelo) ya que influyen en su óptimo desarrollo en todo su ciclo fenológico (Páez et al., 2000).
En términos generales, el clima es el principal factor ecológico que influye en los cambios de la fenología y la composición vegetal, también influyen notablemente las condiciones hídricas como los excesos de agua (condiciones de anegación) y el déficit de agua (Gliessman, 2002). Siendo el estrés producido por la falta de agua, el que provoca modificaciones en sus propiedades mecánicas, es decir cambios incluyen una compleja red de señales químicas y físicas que interactúan entre los sistemas planta-planta y planta-medio ambiente (Caicedo-Lopez et al., 2020)
La sequía demanda mejorar la gestión eficaz del agua para la producción de cultivos en zonas de escases de agua. En este sentido se han buscado aumentar la productividad de los cultivos con técnicas encaminadas a realizar un uso eficiente y eficaz del agua. Estas técnicas involucran combinación entre medidas biológicas como manipulación de semillas, edad de plántulas, y de ingeniería como son sistemas de riego, y almacenamiento de agua de lluvia (Ali & Talukder, 2008). En América Latina se prevé que los sistemas de riego aumenten hasta un 119% entre los años 2000-2080 (Mancosu et al., 2015). Con el propósito de suplir la demanda hídrica en periodos de escases se realizan obras destinadas a almacenar agua en periodos de lluvia (Food and Agriculture Organizatión [FAO], 2013).
La presente investigación tiene por objetivo evaluar el efecto de aplicación de diferentes láminas de riego sobre las etapas fenológicas del cultivo de tomate (Solanum lycopersicum L.) en el municipio de Totogalpa, corredor seco de Nicaragua.
MATERIALES Y MéTODOS
Ubicación del area de estudio
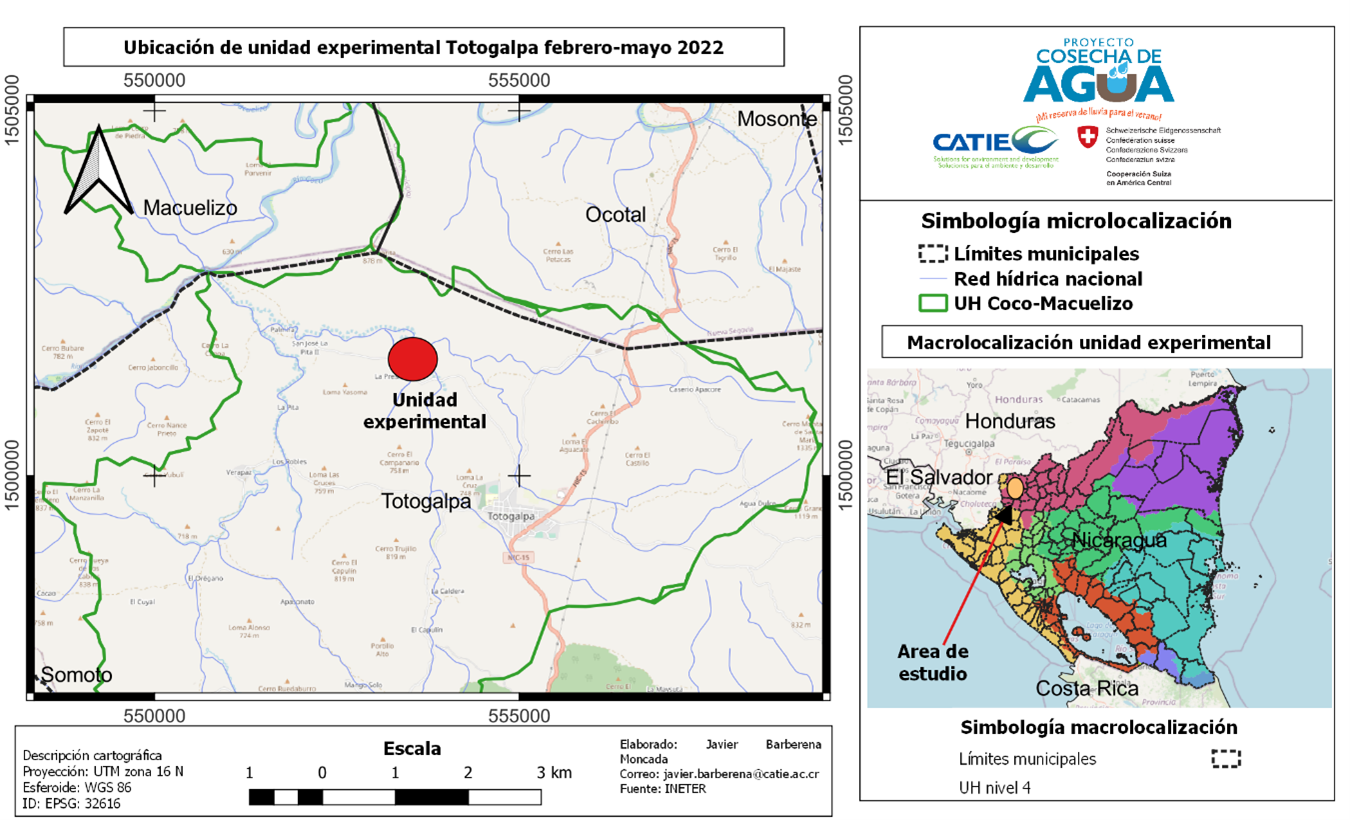
El estudio se realizó en la Finca La Presa Comarca San José de Palmira, con coordenadas 13°35'1.63" latitud norte y 86°30'23.85" longitud oeste, que corresponde al municipio de Totogalpa, departamento de Madriz, Nicaragua (Figura 1).
Totogalpa es un municipio con una extensión aproximada de 133.1 km2, una elevación entre los 660 m.s.n.m con población de 16822 habitantes hasta el año 2021 (Instituto Nacional de Información de Desarrollo [INIDE], 2022). Se caracteriza por presentar temperaturas máximas de 38.6°C, mínimas de 16.1°C, datos registrados mediante estación meteorológica Davis.
Figura 1
Ubicación de área de estudio.
Diseño Experimental
Se estableció unidad experimental en los meses de febrero a mayo de 2021, esta contó de seis surcos de 15 m de largo y 0.8 m de ancho, con una separación entre surcos de 0.8m, separación entre plantas de 0.5m, con diseño completo al azar (DCA), con tres tratamientos y tres réplicas completamente aleatorizado, cada tratamiento cuenta con 30 plantas. Los tratamientos fueron determinados en función de suplir pérdidas por evapotranspiración de la planta, utilizando la ecuación aplicada por (J. Álvarez-Herrera et al., 2010; J. G. Álvarez-Herrera et al., 2012; Balaguera et al., 2008):
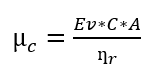 Ecuación 1
Ecuación 1
Donde: µc: cantidad de agua a aplicar (mm). Ev: evaporación en mm. C: coeficiente multiplicador de la evapotranspiración (0.6, 0.8, 1.0). A: área de la unidad experimental (m) ƞr: eficiencia del riego (0,7) (Allen et al., 2006).
Los datos de evaporación fueron calculados mediante CropWat (versión 8.0), los parámetros correspondiente al cultivo se tomaron de Allen et al., (2006) las condiciones edáficas y climáticas de registros de INETER.
Tratamientos a evaluar:
- T1: suplir las pérdidas por evapotranspiración de la planta de tomate en 60%, esto generó un volumen de agua de 2.4 a 3.5 L planta/día.
- T2: suplir las pérdidas por evapotranspiración de la planta de tomate en 80%, generando un volumen de agua de 3.2 a 4.7 L planta/día.
- T3: suplir las pérdidas por evapotranspiración de la planta de tomate en 100%, generando un volumen de agua de 4.1 a 5.9 L planta/día.
Modelo de crecimiento logístico
El modelo utilizado es una ecuación matemática simple que se deriva de una función logística en forma de S, modelando la tasa de crecimiento de una población ya sea en biología o en ecología. El modelo logístico ha sido utilizado por Wardhani & Kusumastuti (2013), con la siguiente ecuación:
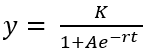 ecuación 2
ecuación 2
Donde:
y: largo de tallo (cm), diámetro de tallo (cm) o número de hojas. t: tiempo (semana). K= capacidad de carga o crecimiento máximo (cm). A= constante. r= tasa de crecimiento (cm)
Se realizó el ajuste de curvas en InfoStat, esto permitió encontrar los valores de K, A y r de la Ecuación 2.
En cada uno de los tratamientos se realizaron mediciones una vez por semana de los factores:
- Crecimiento: largo y ancho de tallo principal en cm, número de hojas.
- Reproductivo: número de flores, número de frutos, diámetro polar y ecuatorial del fruto en cm, peso del fruto en gr.
Tiempos termales
El modelo para calcular el tiempo termal (GDD) se realizó mediante la fórmula propuesta por Pathak & Stoddard (2018):
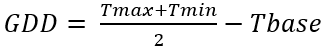 ecuación 3
ecuación 3
Donde:
GDD: Grados días de crecimiento. Tmax: temperaturas máximas (°C). Tmin: temperaturas mínimas (°C). Tbase: temperatura base (°C)
Se realizaron los cálculos diarios desde el trasplante de las plantas de tomates en el campo hasta la cosecha de los frutos. La temperatura base utilizada fue de 10°C, esto debido que las temperaturas medias no excedieron los 30°C (Pathak & Stoddard, 2018) . La acumulación del tiempo termal se realizó mediante la suma de los valores diarios desde el trasplante hasta la cosecha.
Análisis estadísticos
Análisis de varianza más la comparación de Tukey-Kramer (α = 0,05), para determinar si existen diferencias significativas en las medias de las variables de crecimiento y reproductiva del tomate entre los tratamientos utilizados. Se utilizó InfoStat para generar los análisis estadísticos de varianza, Tukey-Kramer, Tukey, los gráficos fueron realizados en SigmaPlot 11.0.
Resultados y discusión
Efecto del estrés hídrico sobre las etapas fenológicas del cultivo de tomate (Solanum lycopersicum L.) variedad INTA JL5 en el período febrero-mayo 2022.
El comportamiento en crecimiento del tomate durante el ciclo del cultivo comprende dos fases: vegetativa con durarión de 35 DDT y una reproductiva de duración de 42 DDT (Figura 2). La primer fase o vegetativa con dos etapas de crecimiento una de establecimiento del cultivo a 7 DDT y 34.44 GDD y la segunda etapa de desarrollo y crecimiento vegetativo hasta 34 DDT y 502.03 GDD.
En la fase reproductiva se distinguen tres etapas, primera floración y cuajo del fruto 42 DDT y 619.6 GDD, segunda etapa desarrollo del fruto hasta los 56 DDT y 857.92 GDD, y una tercera etapa de cosecha 77 DDT y 1201.37 GDD (Figura 2). En la Figura 3 se indica curvas de desarrollos uniformes para todos los tratamientos en las etapas de crecimiento: largo de tallo principal y ancho de tallo principal.
Figura 2
Desarrollo fenológico del cultivo de tomate INTA JL5.

Nota: DDT: días después de siembra. GDD: grados termales.
La variable de crecimiento largo de tallo principal (Tabla 2) presenta diferencias entre los tratamientos (p < 0.05). Todos los tratamientos presentan crecimiento indeterminado en todo el ciclo del cultivo de tomate.
El crecimiento del tallo principal sigue una curva logística. El tallo presenta crecimiento exponencial hasta los 42 DDT, con una acumulación de 619.6 GDD, con crecimiento entre 42-44 cm (Figura 3). La tasa de crecimiento oscila entre 0.49 a 0.6 cm.GDD-1 (Tabla 1). En las etapas de desarrollo del fruto, el crecimiento en largo del tallo es constante en los tres tratamientos hasta la etapa de cosecha, este comportamiento es característicos del cultivo de tomate (Enriquez-Reyes et al., 2003; Juárez-Maldonado et al., 2017).
El T3 es el que mayor media de largo presenta, seguido de T2 y T1. Estudios previos condicionan el crecimiento del tallo a la densidad de plantas por unidad de área (Ayarna et al., 2021). Esto, a diferencia de otros estudios en los que indican que las discrepancias se dan por el origen del material genético, manejo agronómico del cultivo, condiciones climáticas y a estrés hídrico en las plantas (Rodríguez et al., 2013).
Tabla 1
Resultado de modelo de crecimiento lógistico.
| Variable de respuesta | Tratamiento | A | IC 95% | K | IC, 95% | r | IC 95% |
|---|---|---|---|---|---|---|---|
| Largo de tallo (cm) | T1 | 6.29 a | 4.41 - 7.13 | 49.44 b | 48.48 - 50.26 | 0.54 a | 0.47 - 0.58 |
| T2 | 6.55 a | 4.79 - 7.55 | 50.09 b | 49.00 - 50.71 | 0.60 a | 0.51 - 0.66 | |
| T3 | 5.85 a | 4.92 - 6.55 | 55.77 a | 54.04 - 57.03 | 0.49 a | 0.45 - 0.54 | |
| Diámetro de tallo (cm) | T1 | 4.81 a | 2.76 - 6.01 | 1.20 b | 1.15 - 1.26 | 0.64 a | 0.49 - 0.80 |
| T2 | 6.68 a | -5.14 - 8.73 | 1.29 a, b | 1.25 - 1.33 | 0.81 a | 0.39 - 1.04 | |
| T3 | 7.08 a | 2.33 - 8.73 | 1.40 a | 1.31 - 1.40 | 0.78 a | 0.57 - 0.98 | |
| Número de hojas | T1 | 7.49 a | 1.74 - 10.02 | 128.90 | 107.00 - 148.90 | 0.41 b | 0.17 - 0.53 |
| T2 | 20.20 a | -14.35 - 27.04 | 171.43 a | 157.40 - 180.50 | 0.82 a | 0.48 - 0.95 | |
| T3 | 16.63 a | 3.49 - 21.64 | 184.38 a | 178.30 - 189.10 | 0.77 a | 0.55 - 0.86 |
Nora: K= capacidad de carga o crecimiento máximo (cm). A= constante. r= tasa de crecimiento (cm).
La variable de crecimiento diámetro del tallo presenta una curva de crecimiento logística al igual que el largo de tallo principal. Con crecimiento exponencial desde los 7 DDT hasta los 42 DDT, requieriendo 619 GDD-1 para aumentar el diametro (Figura 3). El crecimiento disminuye en las etapas de desarrollo del fruto y cosecha. El diámetro entre los tratamientos fue distinto (p < 0.05), en el cual, T3 presenta mayor media con 0.99 cm y mayor longitud 1.4 cm (ver Tabla 1). Seguido de T2 con media de 0.95 cm y mayor diámetro de 1.29 cm. El que menor diámetro presenta es T1 con media de 0.90 cm y mayor diámetro de 1.2 cm. Los diámetros de tallo de tomate del estudio son superiores a los reportados por Calero H. et al (2019), quien reporta diámetros máximos de 0.6 cm.
Las diferencias entre los diámetros causado por el área del parénquima en el tallo, al presentar mayor área de asimilados favorecen el crecimiento debido a que aumentan el flujo en el xilema y floema hacia los órganos reproductivos del fruto (Ortega-Martínez et al., 2010). De esta manera, los tallos se contraen durante el día por la incorporación de agua en la corriente de evapotranspiración, esto se da en el xilema y floema, y durante la noche se expanden por rehidratación del floema (Gallardo et al., 2006). Por tanto, la pérdida de agua que se produce en los tratamientos con menor agua disponible en la planta (T1 y T2) genera estrés hídrico, debido a las contracciones en el tallo, ocasionando disminución de los conductos del xilema. Como resultado hay una disminución en el crecimiento, en el diámetro del tallo de toda la planta, con poca disponibilidad de agua en comparación con las plantas con mayor agua disponible (T3) (Goldhamer & Fereres, 2001).
El número de hojas presenta mayor generación hasta los 42 DDT y 619 GDD. El número de hojas/planta entre los tratamientos fue distinto (p < 0.05). Los mejores promedios son T3 con 118 hojas/planta y T2 con 110 hojas/planta. Por consiguiente, la mayor producción de hojas/planta, T3 con 184 y T2 con 171. T1 con los menores promedios en número medio de hojas/planta con 70. La producción de hojas en una planta, es asociado a la densidad de siembra, por la competencia por agua y radiación, siendo este último importante por la actividad fotosintética que se dan en las hojas (Barraza et al., 2004).
El factor fundamental es el cierre parcial de estomas para reducir la transpiración y mantener el potencial hídrico de las hojas. Este proceso ocurre como respuesta de la planta cuando existe demanda de agua (Zegbe et al., 2007). El cierre parcial de los estomas, produce una reducción en el tamaño de las hojas debido que se reducen las células estomáticas (Sam et al., 2000). Hay que destacar que, en este estudio no se midió el tamaño de hojas, además, el cierre parcial que ocasiona el estrés hídrico provoca disminución en la conductancia estomática, tasa de fotosíntesis y aumento en la tasa de respiración nocturna. Estos procesos causan reducción de contenido de todas las formas de clorofila en todas las etapas del desarrollo en las plantas de tomate (Yuan et al., 2015).
La inflorescencia en el cultivo de tomate inicia a los 42 DDT con 619 GDD, en etapa de floración y cuajo, esta se extiende hasta el fin del ciclo del cultivo 77 DDT y 1201 GDD. Al igual que las hojas, la cantidad de flores/planta entre los tratamientos varió (p < 0.05), donde los mayores promedios de flores/planta son T3 (18) y T2 (17) y el menor promedio T1 (14). Este comportamiento se debe a que la planta de tomate requiere mayor agua en los órganos reproductivos. Un déficit hídrico no solo reduce el número de flores por planta, también reduce la tasa de aparición de primeras flores (Vijitha & Mahendran, 2010), como ocurrió con el T1 que se retrasó 7 días en comparación con T3 y T2.
Un efecto que causa el estrés hídrico en esta etapa de reproducción y que no fue medido en este estudio es el aborto de flores, reducción del número de óvulos por flor, esto ocasiona una disminución en el número de frutos por planta de tomate (Sibomana et al., 2013). Esta etapa también es afectada por las altas temperaturas y humedad relativa máximas, el daño se produce directamente en la formación de polen, endotelio, epidermis, reducción de los estromas (Sato et al., 2002). También se han reportado que pueden ocurrir caída de capullos, desarrollo anormal de las flores, baja producción de polen (Hazra et al., 2007).
La infrutescencia en las plantas de tomate se da 77 DDT y 1201 GDD. Con diferencias entre número medio de frutos/planta entre los tratamientos (p < 0.05), T3 presenta mayor media de frutos/planta (6.01), seguido de T1 (4.72) y el menor T2 (4.01). Aunque se han reportado valores de 1200 GDD para maduración de cultivos de tomate con una duración de 115-140 DDT con siembra en el mes de marzo, los tiempos de maduración se reduce en 40 días cuando aumenta la temperatura máximas y mínimas. Esto produce que la acumulación de calor en la planta produzca una reducción en los tiempos de maduración de la fruta de tomate (Pathak & Stoddard, 2018). Guaymasí (2015) en su estudio con dos híbridos de tomate, registró etapas de fructificación entre 60-79 DDT, y acumulación de calor de 918 GDD para alcanzar la última fase para cosecha. Ahora bien, Aikman (1996) considera que acumulación de 840 GDD es suficiente para generar maduración de los frutos de tomate. Igualmente Adams et al (2001) estima que los frutos de tomate maduran después, 812 GDD. Estos datos son menores que los resultados presentados en el estudio, por lo que hay que considerar las condiciones climáticas y variedad de tomate utilizada en los experimentos.
Otro factor que no se consideró en los estudios antes mencionados, fue la dependencia que tiene el desarrollo de fruto a la gran cantidad de agua que tiene que estar disponible en la planta de tomate (Ho, 1996; Sibomana et al., 2013). El déficit hídrico causado por las temperaturas es el aborto prematuro de los frutos (Van Der Ploeg & Heuvelink, 2005). Otros síntoma que causan las temperaturas es rajado de frutos y reducción en peso fresco (Rodriguez-Ortega et al., 2017). Se cree que estos síntomas se dan durante las horas de medio día que es cuando ocurren las mayores temperaturas (Pearce et al., 1993a, 1993b).
Es de suma importancia tener monitoreo constante de las variables y hacer estudios en otras variedades de tomate, esto debido a que los resultados de las proyecciones climáticas presentan una tendencia de aumento de temperatura de hasta los 2ºC al 2050 (Hannah et al., 2017).
Los diámetro polar y ecuatorial del fruto variaron entre tratamientos (p < 0.05). En ellos T3 presentó mayor diámetro, con una media en su diámetro polar de 5.94 cm y ecuatorial 5.84 cm, seguido de T2 con media de 4.8 cm y 4.34 cm de diámetro polar y ecuatorial respectivamente. En el cual, T3 con menor tamaño registra diámetro polares y ecuatoriales de 4.2 cm y 3.69 cm respectivamente. Los diámetros obtenidos en T3 concuerdan con los resultados presentados por Casierra-Posada et al (2007) quien presenta diámetros entre 6-8 cm. Casierra-Posada et al (2007) reporta diámetro de 5 cm (polar y ecuatorial), el cual cree que la disminución del crecimiento del fruto ocurre después de los 57 días de la floración; esto debido a pérdidas por transpiración del fruto.
En el estudio de Ardila R. et al (2012), los menores diámetros en frutos coinciden con el mejor rendimiento del presente estudio que es T1, reportando diámetros entre 5.9 a 7.38 cm y 1398 GDD, este estudio alcanza los 1201 GDD. La diferencia de tamaño polar y ecuatorial se asocia a la retención de agua del fruto, que está compuesto en un 90-95% de agua y el restante 5-10% de materia seca (Wang et al., 2011). El estrés hídrico causa que el agua no llene lo suficiente los vasos conductores de agua, cuyo movimiento se realiza mediante el xilema y floema. S estima que un máximo de 90% del agua pasa por el floema y el restante 10% por el xilema (Guichard et al., 2003; L. C. Ho et al., 1987).
Se destaca el hecho que, los frutos difieran en tamaño y peso debido a la acumulación de agua. Frutos con déficit hídrico presentan el mismo contenido de materia seca que los cultivados con buenas condiciones hídricas; esto se debe a que la transpiración que ocurre en el fruto aumente las concentraciones de sólidos suspendido totales producidas por la pérdida de agua en el fruto (Casierra Posada et al., 2007; Van de Wal et al., 2017).
Figura 3
Etapas de crecimiento de plantas de tomate
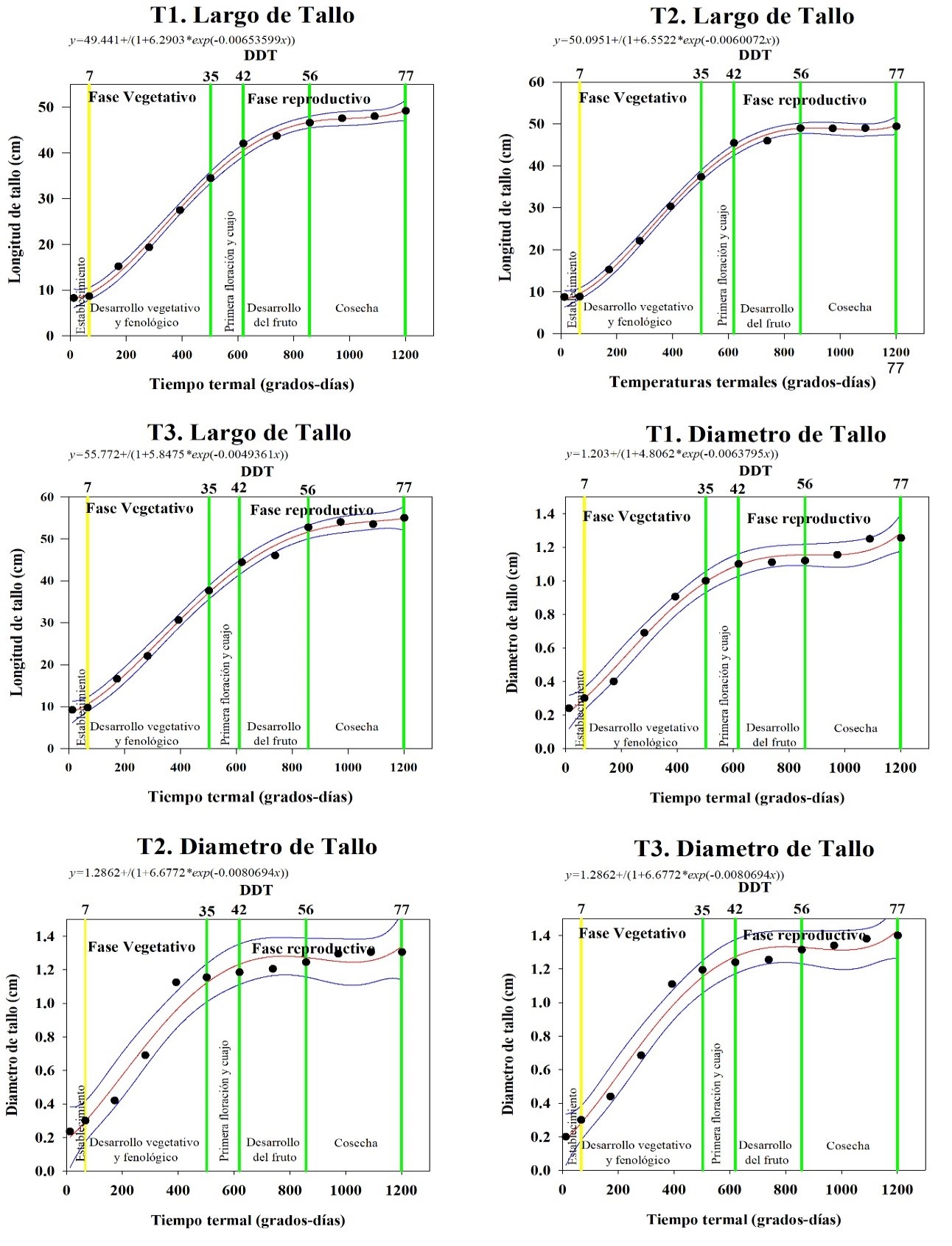
Nota: líneas amarillas y verdes indican las distintas etapas de crecimiento del cultivo.
El rendimiento de tomate/planta fue distintos entre los tratamientos evaluados (p < 0.05). El cual, T3 es el que presenta mayor promedio en el peso fresco de cada fruto (68.4g), seguido T2 (36.5g) y con menor promedio en peso fresco de fruto T1 (22.3g). Extrapolando a rendimientos por hectárea (ha), se tendrían rendimientos en T3 de aproximadamente 8129.4 kg.ha-1, seguido de T2 con rendimiento promedio de 3313.2 kg.ha-1 y T1 con rendimientos medio de 1699.3 kg. ha-1. González Meza (2000) obtuvo resultados de 35.8 ta.ha-1 al suplir en 80% la ET del cultivo de tomate. Aunque difiere de nuestro estudio, durante el experimento se presentó problemas de podredumbre de frutos en el tratamientos de 100% de ET que no permitieron tener mejores resultados.
Ahora bien, López (2016) reporta mayores rendimientos con relación al presente estudio, alcanzando 13500 kg.ha-1 para la variedad de tomate UC-82, experimento realizado en Managua, Nicaragua. Asi mismo, Jiménez-Martínez et al (2010) realizó un estudio en Ciudad Sandino, Nicaragua, con un experimento en cuatro variedades de tomate, obteniendo rendimientos similares a este estudio, cuyos rendimiento se encuentran entre 4406 a 7138 kg.ha-1.
Aunque estos estudios obtienen rendimientos distintos a la presente investigación, es importante señalar la importancia de suplir las necesidades hídricas de la planta de tomate, debido que afecta grandemente el rendimiento del cultivo, puesto el déficit hídrico no solo afecta el crecimiento de la planta, también al número de frutos por planta, tamaño y peso de fruto. Estas afectaciones son producto del desequilibrio que ocurre en la planta, en los procesos de fotosíntesis y respiración en la planta y el fruto (Chaves et al., 2003; Rodriguez-Ortega et al., 2017).
Para mantener un buen rendimiento del cultivo de tomate, las plantas deben de tener el volumen de agua necesario. También la intensidad y frecuencia de estas son claves, debido a que la interrupción del riego en los procesos como la floración y desarrollo del fruto trae consecuencias en el rendimiento (Wang & Xing, 2017). El riego en combinación con técnicas de fertilización, podas de formación aumentan la calidad del fruto y su rendimiento (Bertin & Génard, 2018).
Tabla 2
Análisis de varianza de variables de crecimiento y reproducción en los tres tratamientos de tomate
| Etapa fenológica | Variable | Tratamiento | Media | E.E | p-valor |
|---|---|---|---|---|---|
| Crecimiento | Largo de tallo | T1 | 33.33b | 0.45 | 0.0160 |
| T2 | 34.18b | ||||
| T3 | 35.98A | ||||
| Diámetro de tallo | T1 | 0.90c | 0.01 | 0.0021 | |
| T2 | 0.95b | ||||
| T3 | 0.99a | ||||
| Número de hojas | T1 | 73.15c | 2.75 | 0.0014 | |
| T2 | 110.78a | 0.85 | |||
| T3 | 118.76a | 0.70 | |||
| Reproductiva | Número de flores | T1 | 15.00b | 0.13 | 0.0004 |
| T2 | 17.30a | 0.34 | |||
| T3 | 18.33a | 0.51 | |||
| Número de frutos | T1 | 4.72b | 0.21 | 0.0014 | |
| T2 | 4.01b | ||||
| T3 | 6.01a | ||||
| Diámetro polar de fruto | T1 | 4.20c | 0.11 | < 0.0001 | |
| T2 | 4.80b | ||||
| T3 | 5.94a | ||||
| Diámetro ecuatorial de fruto | T1 | 3.69c | 0.09 | < 0.0001 | |
| T2 | 4.34B | ||||
| T3 | 5.48a | ||||
| Peso de fruto | T1 | 22.36C | 2.43 | < 0.0001 | |
| T2 | 36.57B | ||||
| T3 | 68.43A |
Nota: Medias con igual letra no son significativamente diferentes (p > 0.05).
CONCLUSIONES
Tanto las distintas etapas de crecimiento y reproducción del cultivo de tomate se ven afectadas cuando no se suplen las pérdidas por evapotranspiración. En este sentido se demostró que el crecimiento de sus órganos vegetativos, reproductivos y el rendimiento del fruto se ven disminuidos por la falta de agua, en esto, suplir el 100% las pérdidas diarias por evaporación del cultivo de tomate es indispensable para un desarrollo óptimo.
También, la utilización de GDD como modelo biotérmico resulta ser buen predictor en la dinámica de cultivo de tomate, estableciendo que para el cultivo de tomate en la zona de Totogalpa se necesita 1200 GDD y 77 DDT. Más aún nos indica que las fases vegetativa tiene un crecimiento exponencial hasta los 42 DDT, luego disminuye debido al inicio de la fase de la fruta y llenado de los frutos.
Este modelo de es suma importancia ya que, se pueden hacer simulaciones de proyecciones si hay aumento o disminución de temperaturas y poder predecir qué sucederá con los tiempos fisiológicos del cultivo de tomate, si se reducen o aumentan. El monitoreo de variables climáticas es muy importante para futuros estudios, principalmente la temperatura máximas y mínimas debido a que afectan el crecimiento en todas las etapas de la planta, pero principalmente afecta en la etapa reproductiva siendo las flores y frutos los principales afectados, reduciendo los rendimientos.
REFERENCIAS BIBLIOGRáficas
Aikman, D. P. (1996). A procedure for optimizing carbon dioxide enrichment of a glasshouse tomato crop. Journal of Agricultural and Engineering Research, 63(2), 171–183. https://doi.org/10.1006/jaer.1996.0019
Ali, M. H., & Talukder, M. S. U. (2008). Increasing water productivity in crop production-A synthesis. Agricultural Water Management, 95(11), 1201–1213. https://doi.org/10.1016/j.agwat.2008.06.008
Allen, R. G., Pereira, L. S., Raes, D., & Smith, M. (2006). Evapotranspiración del cultivo. Guías para la determinación de los requerimientos de agua de los cultivos (FAO, Issue Estudio FAO. Riego y Drenaje). FAO. https://www.fao.org/3/x0490s/x0490s00.htm
Álvarez-Herrera, J. G., López, J. L., Balaguera, W. A., Merchán, J. B., & Veloza, J. A. (2012). Láminas de riego y calidad de agua en la solución de problemas de salinidad en tomate (Solanum lycopersicum L.). Revista Colombiana de Ciencias Hortícolas, 5(1), 57–68. https://doi.org/10.17584/rcch.2011v5i1.1253
Álvarez-Herrera, J., H, B.-L., & Chacón, E. (2010). Efecto de la aplicación de diversas láminas y frecuencias de riego en la propagación del romero ( Rosmarinus officinalis L . ) The effect of applying different water levels and irrigation frequencies in. Ingeniería e Investigación, 30(1), 86–90.
Ardila R., G., Fischer, G., & Balaguera López, H. E. (2012). Caracterización del crecimiento del fruto y producción de tres híbridos de tomate (Solanum lycopersicum L.) en tiempo fisiológico bajo invernadero. Revista Colombiana de Ciencias Hortícolas, 5(1), 44–56. https://doi.org/10.17584/rcch.2011v5i1.1252
Ayarna, A. W., Tsukagoshi, S., Nkansah, G. O., & Maeda, K. (2021). Effect of Plant Density on the Yield of Hydroponically Grown Heat-Tolerant Tomato under Summer Temperature Conditions. American Journal of Plant Sciences, 12(06), 901–913. https://doi.org/10.4236/ajps.2021.126060
Balaguera, H. E., Álvarez-Herrera, J., & Rodríguez, J. D. (2008). Efecto del déficit de agua en el trasplante de plántulas de tomate (Solanum lycopersicum L.). Agronomía Colombiana, 26(2), 246–255. http://hdl.handle.net/20.500.12324/34753
Barraza, F. V., Fischer, G., & Cardona, C. E. (2004). Estudio del proceso de crecimiento del cultivo del tomate (Lycopersicon esculentum Mill.) en el Valle del Sinú medio, Colombia. Agronomía Colombiana, 22(1), 81–90. http://www.redalyc.org/articulo.oa?id=180317823011
Bertin, N., & Génard, M. (2018). Tomato quality as in fl uenced by preharvest factors. Scientia Horticulturae, 233, 264–276. https://doi.org/10.1016/j.scienta.2018.01.056
Caicedo-Lopez, L. H., Contreras-Medina, L. M., Gerardo, R., Perez-Matzumoto, A. E., Ruiz-Rueda, A., Helena, L., Contreras-Medina, L. M., Guevara-
Gonzalez, G., Perez-Matzumoto, A. E., Ruiz-Rueda, A., Caicedo-Lopez, L. H., & Contreras-Medina, L. M. (2020). Effects of hydric stress on vibrational frequency patterns of Capsicum annuum plants Effects of hydric stress on vibrational frequency patterns of Capsicum annuum plants. Plant Signaling & Behavior, 15(7). https://doi.org/10.1080/15592324.2020.1770489
Calero H., A., Quintero R., E., Pérez D., Y., Olivera V., D., Peña C., K., Castro L., I., & Jiménez H., J. (2019). Evaluación de microorganismos eficientes en la producción de plántulas de tomate (Solanum lycopersicum L.). Revista de Ciencias Agrícolas, 36(1), 67–78. https://doi.org/10.22267/rcia.193601.99
Calvo-Solano, O. D., Quesada-Hernández, L. E., Hidalgo, H., & Gotlieb, Y. (2018). Impactos de las sequías en el sector agropecuario del Corredor Seco Centroamericano. Agronomía Mesoamericana, 29(3), 695. https://doi.org/10.15517/ma.v29i3.30828
Casierra-Posada, F., Constanza Cardozo, M., & Cárdenas-Hernández, J. F. (2007). Análisis del crecimiento en frutos de tomate (Lycopersicon esculentum Mill .) cultivados bajo invernadero. Agronomía Colombiana, 25(2), 299–305.
Casierra Posada, F., Cardozo, C. M., & Cárdenas-Hernández, J. F. (2007). Análisis del crecimiento en frutos de tomate (Lycopersicon esculentum Mill.) cultivados bajo invernadero. 25(2), 299–305. http://hdl.handle.net/20.500.12324/979
Chaves, M. M., Maroco, J. P., & Pereira, J. S. (2003). Understanding plant responses to drought - From genes to the whole plant. Functional Plant Biology, 30(3), 239–264. https://doi.org/10.1071/FP02076
Duarte-Canales, H. A., & Benavides González, Á. (2020). Riego por goteo en cultivares de tomate (Solanum lycopersicum L.), Universidad Nacional Agraria, Managua, Nicaragua. La Calera, 20(34), 35–40. https://doi.org/10.5377/calera.v20i34.9811
Enriquez-Reyes, S., Alcántar-González, G., Castellanos-Ramos, J., Arjona Suárez, E., González-Eguiarte, D., & Lazcano-Ferrat, I. (2003). Nutrición Mineral Acoplada Al Crecimiento (Numac): Nutrición Con N Para Tomate En Invernadero 3. Evaluación del Modelo. Terra Latinoamericana, 1(2), 185–193. http://redalyc.uaemex.mx/redalyc/html/573/57315595005/57315595005_1.html
FAO. (2013). Captación Y Almacenamiento De Agua De Lluvia. In Santiago de Chile.
Fraga, F. (2020). Corredor seco Centroamericano: Una visión exploratoria sobre el contexto, las razones y el potencial de una estrategia de creación de empleo en Guatemala y Honduras (O. I. del Trabajo (ed.); Primera Ed, Issue 23). www.ifrro.org
Gallardo, M., Thompson, R. B., Valdez, L. C., & Fernández, M. D. (2006). Use of stem diameter variations to detect plant water stress in tomato. Irrigation Science, 24(4), 241–255. https://doi.org/10.1007/s00271-005-0025-5
Gliessman, S. R. (2002). Agroecología: Procesos ecológicos en agricultura sostenible. In Diversidad y estabilidad del agroecosistema (CATIE). CATIE.
Goldhamer, D. A., & Fereres, E. (2001). Irrigation scheduling protocols using continuously recorded trunk diameter measurements. Irrigation Science, 20(3), 115–125. https://doi.org/10.1007/s002710000034
González Meza, A., & Hernández Leos, B. A. (2000). Estimación de las necesidades hídricas del tomate. Terra Latinoamericana, 18(1), 45–50.
Guaymasí, D. V. (2015). Estudio de la respuesta fenológica a través de la determinación del tiempo térmico de las Solanáceas más producidas en el Cinturón Hortícola Platense. In Tesis doctoral. Universidad Nacional de La Plata.
Guichard, S., Bertin, N., Leonardi, C., & Gary, C. (2003). Tomato Fruit quality in relation to water and carbon fluxes. Agronomie, 21(4), 385–392. https://doi.org/10.1051/agro:2001131
Hannah, L., Donatti, C. I., Harvey, C. A., Alfaro, E., Rodriguez, D. A., Bouroncle, C., Castellanos, E., Diaz, F., Fung, E., Hidalgo, H. G., Imbach, P.,
Läderach, P., Landrum, J. P., & Solano, A. L. (2017). Regional modeling of climate change impacts on smallholder agriculture and ecosystems in Central America. Climatic Change, 141(1), 29–45. https://doi.org/10.1007/s10584-016-1867-y
Hazra, P., Samsul, H., Sikder, D., & Peter, K. (2007). Breeding tomato (Lycopersicon esculentum Mill) resistant to high temperature stress. International Journal of Plant Breeding, 1(1), 31–40. http://www.globalsciencebooks.info/Online/GSBOnline/images/0706/IJPB_1(1)/IJPB_1(1)31-40o.pdf
Ho, L. (1996). The mechanism of assimilate partitioning and carbohydrate compartmentation in fruit in relation to the quality and yield of tomato. Journal of Experimental Botany, 47(Special_Issue), 1239–1243. https://doi.org/https://doi.org/10.1093/jxb/47.Special_Issue.1239
Ho, L. C., Grange, R. I., & Picken, A. J. (1987). An analysis of the accumulation of water and dry matter in tomato fruit. Plant, Cell & Environment, 10(2), 157–162. https://doi.org/10.1111/1365-3040.ep11602110
INIDE. (2022). Anuario Estadístico 2020. https://www.inide.gob.ni/docs/Anuarios/Anuario20/Anuario_Estadistico_2020.pdf
IPCC. (2019). El cambio climático y la tierra. 40.
Jiménez-Martínez, E., Gutiérrez Sandoval, W. A., & González Madrigal, C. A. (2010). Evaluacion de cuatro variedades de tomate industrial (Lycopersicum esculentum, Mill) en el rendimiento y tolerancia al complejo mosca blanca (Bemisia tabaci Gennadius)-Geminivirus. La Calera, 10(15), 5–15.
Jiménez-Martínez, E., Ríos-Peralta, H. T., & Somarriba-Moncada, O. A. (2015). Evaluación de productos botánicos para manejo de mosca blanca (Bemisia tabaci Gennadius) y pulga del tomate (Halticu ssp) en el cultivo de tomate (Solanum lycopersicum Mill.), en Nicaragua. La Calera, 15(25), 63–69.
Juárez-Maldonado, A., De Alba Romenus, K., Zermeño González, A., & Benavides Mendoza, A. (2017). Análisis de crecimiento del cultivo de tomate en invernadero. Revista Mexicana de Ciencias Agrícolas, 6(5), 943–954. https://doi.org/10.29312/remexca.v6i5.589
López Ubeda, P. E., & Coleman Beer, E. R. (2016). Efecto de tres láminas de riego por goteo y tres distancias de siembra en el cultivo de tomate (Solanum lycopersicum L.) Cv. UC-82, Universidad Nacional Agraria, Managua [Universidad Nacional Agraria]. chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/viewer.html?pdfurl=https%3A%2F%2Frepositorio.una.edu.ni%2F3415%2F1%2Ftnf06l864.pdf&clen=1917390&chunk=true
Magaña, V. O., Amador, J. A., & Medina, S. (1999). The Midsummer Drought over Mexico and Central America. Journal of Climate, 12(6), 1577–1588. https://doi.org/10.1175/1520-0442(1999)012<1577:TMDOMA>2.0.CO;2
Mancosu, N., Snyder, R. L., Kyriakakis, G., & Spano, D. (2015). Water scarcity and future challenges for food production. Water (Switzerland), 7(3), 975–992. https://doi.org/10.3390/w7030975
Ortega-Martínez, L. D., Sánchez-Olarte, J., Ocampo-Mendoza, J., Sandoval-Castro, E., Salcido-Ramos, B. A., & Manzo-Ramos, F. (2010). Efecto de diferentes sustratos en crecimiento y rendimiento de tomate (Lycopersicum esculentum Mill) bajo condiciones de invernadero. / Effect of different substrates on the growth and yield of tomato (Lycopersicum esculentum Mill) under greenhouse condit. Ra Ximhai Revista de Sociedad, Cultura y Desarrollo Sustentable, 6(3), 339–346. http://www.redalyc.org/articulo.oa?id=46116015002
Páez, A., Paz, V., & López, J. . (2000). Crecimiento y respuestas fisiológicas de plantas de tomate cv . Río Grande en la época mayo-julio. Efecto del sombreado. Fac.Agron, 17(2), 173–184. https://produccioncientificaluz.org/index.php/agronomia/article/view/26349
Pathak, T. B., & Stoddard, C. S. (2018). Climate change effects on the processing tomato growing season in California using growing degree day model. Modeling Earth Systems and Environment, 4(2), 765–775. https://doi.org/10.1007/s40808-018-0460-y
Pearce, B. D., Grange, R. I., & Hardwick, K. (1993a). The growth of young tomato fruit. I. Effects of temperature and irradiance on fruit grown in controlled environments. Journal of Horticultural Science, 68(1), 1–11. https://doi.org/10.1080/00221589.1993.11516322
Pearce, B. D., Grange, R. I., & Hardwick, K. (1993b). The growth of young tomato fruit. II. Environmental influences on glasshouse crops grown in rockwool or nutrient film. Journal of Horticultural Science, 68(1), 13–23. https://doi.org/10.1080/00221589.1993.11516323
Rodriguez-Ortega, W., Martinez, V., Rivero, R. M., Camara-Zapata, J., Mestre, T., & Garcia-Sanchez, F. (2017). Use of a smart irrigation system to study the effects of irrigation management on the agronomic and physiological responses of tomato plants grown under different temperatures regimes. Agricultural Water Management, 183, 158–168. https://doi.org/10.1016/j.agwat.2016.07.014
Rodríguez Díaz, E., Salcedo Pérez, Eduardo Rodríguez Macias, Ramón González Eguiarte, D. R., & Mena Munguía, S. (2013). Reúso del tezontle: efecto en sus características físicas y en la producción de tomate (Licopersicom esculentum Mill). Terra Latinoamericana, 31(4), 275–284. http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0187-57792013000500275&lng=es&nrm=iso&tlng=es
Sam, O., Jérez, E., Dell’amico, J., & Ruiz-Sanchez, M. C. (2000). Water Stress Changes and Tomato Anatomy Leaf Epidermes. In Biologia Plantarum (Vol. 43, Issue 2, pp. 275–277). https://doi.org/doi:10.1023/a:1002716629802
Sato, S., Peet, M. M., & Thomas, J. F. (2002). Determining critical pre- and post-anthesis periods and physiological processes in Lycopersicon esculentum Mill. exposed to moderately elevated temperatures. Journal of Experimental Botany, 53(371), 1187–1195. https://doi.org/10.1093/jexbot/53.371.1187
Sibomana, I. C., Aguyoh, J. N., & Opiyo, A. M. (2013). Water stress affects growth and yield of container grown tomato ( Lycopersicon esculentum Mill ) plants. Global Journal of Bio-Science and Biotecnology, 2(4), 461–466.
Van de Wal, B. A. E., Windt, C. W., Leroux, O., & Steppe, K. (2017). Heat girdling does not affect xylem integrity : an in vivo magnetic resonance imaging study in the tomato peduncle. New Phytologist, 215(2), 558–568. https://doi.org/10.1111/nph.14610
Van Der Ploeg, A., & Heuvelink, E. (2005). Influence of sub-optimal temperature on tomato growth and yield: A review. Journal of Horticultural Science and Biotechnology, 80(6), 652–659. https://doi.org/10.1080/14620316.2005.11511994
Van Der Zee Arias, A., Van Der Zee, J., Meyrat, A., Poveda, C., & Picado, L. (2012). Estudio de caracterización del Corredor Seco Centroamericano: Vol. I (FAO). https://reliefweb.int/sites/reliefweb.int/files/resources/tomo_i_corredor_seco.pdf
Vijitha, R., & Mahendran, S. (2010). Effect of moisture stress at different growth stages of tomato plant (Lycopersicon esculentum Mill.) on yield and quality of fruits. Journal of Science of the University of Kelaniya Sri Lanka, 5, 1–11. https://doi.org/http://doi.org/10.4038/josuk.v5i0.4086
Wang, F., Kang, S., Du, T., Li, F., & Qiu, R. (2011). Determination of comprehensive quality index for tomato and its response to different irrigation treatments. Agricultural Water Management, 98(8), 1228–1238. https://doi.org/10.1016/j.agwat.2011.03.004
Wang, X., & Xing, Y. (2017). Evaluation of the effects of irrigation and fertilization on tomato fruit yield and quality : a principal component analysis. Scientific Reports, 7(350), 1–13. https://doi.org/10.1038/s41598-017-00373-8
Wardhani, W. S., & Kusumastuti, P. (2013). DESCRIBING THE HEIGHT GROWTH OF CORN USING LOGISTIC AND GOMPERTZ MODEL. AGRIVITA, 35(3), 237–241. https://doi.org/http://dx.doi.org/10.17503/Agrivita-2013-35-3-p237-241
Yuan, X. K., Yang, Z. Q., Li, Y. X., Liu, Q., & Han, W. (2015). Effects of different levels of water stress on leaf photosynthetic characteristics and antioxidant enzyme activities of greenhouse tomato . 54(1), 28–39. https://doi.org/doi:10.1007/s11099-015-0122-5
Zegbe, J. A., Behboudian, M. H., & Clothier, B. E. (2007). Respuesta del tomate para proceso al reigo parcial de la raíz. Terra Latinoamericana, 25(2395–8030), 61–67.
© 2023 Revista Científica de FAREM-Estelí.
![]() Este trabajo está licenciado bajo una Licencia Internacional Creative Commons 4.0 Atribución-NoComercial-CompartirIgual.
Este trabajo está licenciado bajo una Licencia Internacional Creative Commons 4.0 Atribución-NoComercial-CompartirIgual.